
Adeno-Associated Viral (AAV) Vector Guide
Adeno-Associated Viruses (AAV) are small, non-enveloped viruses containing a small, linear single-stranded DNA (ssDNA) genome. Non-enveloped viruses do not have a lipid bilayer surrounding the viral particle and are only composed of a protein capsid and the viral genetic material inside.
The AAV genome is ~4.7 kb long and consists of two open reading frames, Rep and Cap, flanked by two 145 bp inverted terminal repeats (ITRs). The Rep region encodes four non-structural replication proteins (Rep78, Rep68, Rep52, and Rep40), which are involved in genome replication and viral packaging. The Cap region encodes three structural capsid proteins or viral proteins (VP1, VP2 and VP3) that form the icosahedral capsid of the virus. The ITR sequences serve as origins of replication and also act as signals for packaging.

AAVs are part of the Dependoparvovirus genus, included within the Parvoviridae family. They can infect dividing and non-dividing cells; however, they are replication deficient and depend on other viruses such as adenovirus or herpes simplex virus to replicate in the host cell and complete the lytic cycle. AAVs are therefore non-pathogenic and generally do not trigger a strong immune response. These features have made AAVs the preferred choice to be used as viral vectors for gene therapy applications.
This guide contains many viral vector-specific terms and acronyms, so if you're new to viral vectors or simply need a refresher, we've included a glossary at the end!
Adeno-Associated Virus versus Adenovirus
Adenoviruses (AdV) are part of the Adenoviridae family, are medium-sized, and contain a double-stranded DNA genome. Adeno-associated viruses (AAV), however, are small single-stranded DNA viruses that belong to the Parvoviridae family. AAVs were discovered as a contaminant of adenovirus preparations, which is how they got their name, but they are not related. AAVs require the presence of adenoviral genes E1, E4, E2a and VA for replication.
For more information about adenovirus, read our Adenoviral Vector Guide.
Recombinant AAV Vectors
Wild-type AAVs have been modified by researchers to create recombinant adeno-associated viral (rAAV) vectors in order to deliver genetic cargo into cells. Recombinant AAV constructs are designed by replacing the Rep and Cap regions between the two ITRs with a gene expression cassette that typically contains the transgene of interest. The size of the genetic cargo that can be included between the ITRs is limited by the physical space available inside of the small AAV capsid and directly correlates to the size of the DNA removed from the viral genome (~4.7 kb). For rAAV vector production, the Rep and Cap regions are supplied in trans, via a different plasmid.
In order to produce rAAV vectors, three plasmids are needed:
- Transfer plasmid (also known as cis plasmid) — containing the transgene of interest between the two ITRs.
- Packaging plasmid (also known as Rep/Cap plasmid) — containing the Rep and Cap regions necessary for the production and assembly of viral capsids.
- Helper plasmid — containing adenoviral genes not present in the AAV genome required for viral replication (E4, E2a and VA). These are sometimes combined within the packaging plasmid.
For a summary of all AAV plasmid components, see the AAV Plasmid Elements table.

AAV Vector Production
Cloning
Cloning your transgene, gRNA, or shRNA of interest into the transfer plasmid can be done with most standard cloning methods, including restriction enzyme, Gibson Assembly, or Gateway. Some transfer plasmids may have limited restriction sites or may only be compatible with certain cloning methods (such as a Gateway destination vector), so be sure to confirm your chosen plasmid is compatible with cloning methods available in your lab.
When cloning your plasmids, be sure to use recombination deficient bacterial strains, such as NEB Stable cells. These strains reduce the frequency of homologous recombination of unstable regions, like the ITRs found in transfer plasmids. This will ensure that the repeats will be maintained and often results in a greater yield of DNA. However, if the plasmid contains a Gateway cassette containing the ccdB gene, a ccdB-resistant strain is necessary.
For more information on cloning and working with plasmids, visit Addgene’s Molecular Biology Reference.
Production
The three production plasmids (transfer, packaging, and helper plasmid) are transfected into cells which then act as in vitro factories for the new viral particles in the laboratory. The cell line used for rAAV vector production needs to express the adenovirus E1 gene, which is crucial for efficient AAV replication as it activates the necessary cellular machinery to produce large amounts of rAAV vectors. The E1 gene is not included in the helper plasmid and instead provided by the chosen cell line to ensure safety and prevent the potential formation of replication-competent viruses. While AAV themselves are replication incompetent, there is a rare possibility that recombination between the transfected plasmids could lead to the production of replication-competent viruses that would be able to infect cells and replicate autonomously.
Human embryonic kidney 293T (HEK293T) cells are the most common platform to produce AAV vectors. Additional production systems based on other human cell lines, such as HeLa and A549 cells, and even insect Sf9 cells, have also been developed. The newly formed viral particles produced in HEK293T cells are then extracted and purified for their use as viral vectors to deliver genes to cells in in vivo models.
For more detailed protocols, see Addgene’s Viral Vector Protocols.

Genome Integration
Recombinant AAVs are generally considered to not integrate into the host genome. When wild-type AAVs enter a host cell, they uncoat in the nucleus and present as linear DNA molecules. In the absence of helper viruses, wild-type AAVs cannot enter the lytic cycle, and instead establish latent infections by integrating into the host genome at a specific site, the AAVS1 site on human chromosome 19. This integration is mediated by proteins Rep78 and Rep68, which can bind to specific Rep-binding elements present in the AAV ITRs and the AAVS1 site.
Since the Rep region is removed from the rAAV genome for viral vector production, rAAV vectors do not carry the genes to express Rep78 and Rep 68 proteins and are therefore unable to integrate in the host’s genome. Instead, the linear rAAV DNA is typically processed into a double-stranded circular episome which can concatemerize, producing high molecular weight structures that are maintained extrachromosomally. Random integration of rAAV genomes in non-homologous sites of the host genome can occur, although at a low frequency which has been estimated to range from 0.01% to 3% depending on cell type and rAAV vector concentration. Nonetheless, the majority of rAAV DNA is thought to be maintained in episomes or concatemers. This avoids the effects local chromatin structures in the host genome can have on transgene expression and the expression of neighboring genes. rAAV episomes can develop chromatin-like organization and persist in non-dividing cells for a period of years without damaging the host cell. Episomal stability allows for long-term transgene expression in non-dividing cells while reducing the risk of immune activation, both key advantages of rAAV vectors.
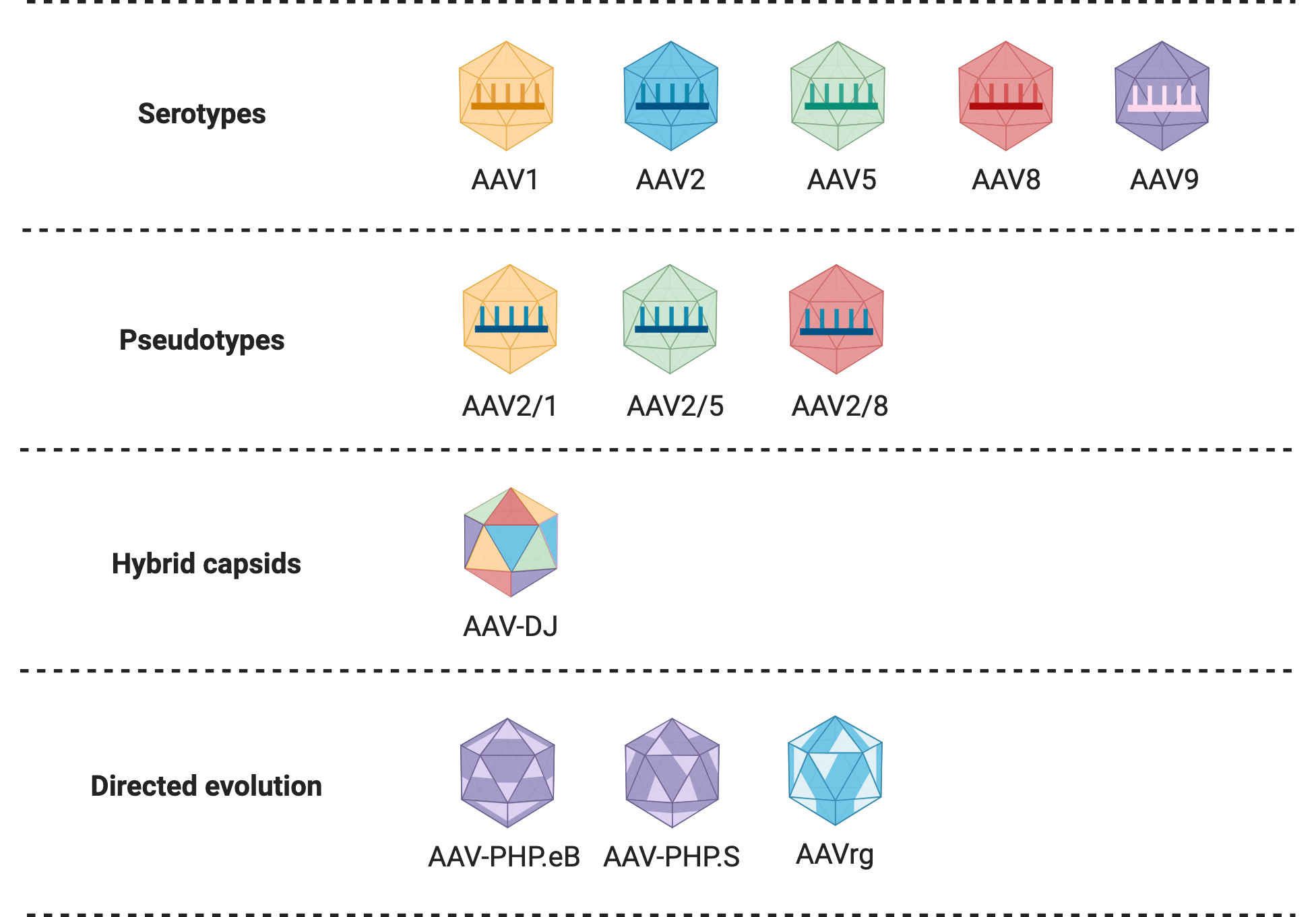
AAV Types
Serotypes
AAV serotypes are AAV variants that differ in their tropism — that is, the types of cells they infect. Serotypes are determined by naturally occurring differences in the variable regions of the virus capsid. Thirteen human and primate serotypes of AAV have thus far been identified, and it is likely that this number will continue to increase. This diversity makes rAAV vectors a very useful system for preferentially transducing specific cell types. The serotype of an rAAV vector is determined by the packaging plasmid used during production, as it carries the Cap region responsible for the production of capsid proteins. AAV2 is the most commonly used serotype for rAAV vector construction, as it was the first to be well-characterized and extensively studied, and has a broad tissue tropism. Table 1 gives a summary of the tropism of AAV serotypes, indicating different serotype(s) for transduction of a given organ.
| Tissue | Serotype |
|---|---|
| Central nervous system | AAV1, AAV2, AAV5, AAV8, AAV9, AAV11, AAVrg, PHP.eB |
| Heart | AAV1, AAV8, AAV9 |
| Kidney | AAV8 |
| Liver | AAV8, AAV9 |
| Lung | AAV6 |
| Pancreas | AAV2, AAV8 |
| Photoreceptor cells | AAV2, AAV5, AAV8 |
| Skeletal muscle | AAV1, AAV8, AAV9 |
Table 1: Summary of tissue tropism displayed by different AAV serotypes (AAV1–11) and engineered variants (AAVrg, PHP.eB).
Pseudotypes
Pseudotyping is the process of producing viral vectors in combination with foreign capsid proteins. Researchers have further refined the tropism of rAAV vectors by mixing capsids and genomes from different viral serotypes. Pseudotypes are generally denoted using two numbers separated by a slash (e.g. AAV2/5), where the first number indicates the serotype of the genome and the second number the serotype of the capsid. Pseudotyped vectors can improve transduction efficiency and alter tropism. For example, AAV2/5, targets neurons that are not efficiently transduced by AAV2, and is distributed more widely in the brain. Many of these hybrid viral vectors have been well characterized and are often preferred over standard rAAV vectors for in vivo applications.
Other Engineered Variants
Hybrid capsids
Hybrid capsids are engineered capsids derived from multiple different serotypes, which can further alter transduction efficiency and viral tropism. One common example is AAV-DJ, which contains a hybrid capsid derived from eight different serotypes. AAV-DJ displays a higher transduction efficiency in vitro than any wild-type serotype, and a very high infectivity across a broad range of cell types in vivo. AAV-DJ has been optimized further for specific cell and tissue types, like AAV-DJ8, which displays enhanced brain uptake.
Systemic capsids
Another strategy to fine-tune tropism towards specific cell types of interest has been to create and test capsid libraries with random mutations or insertions in the Cap region. Several methods have been developed and successfully employed to engineer new rAAV capsid variants, such as systemic capsids. A systemic capsid is a capsid that has been engineered to have enhanced function after intravenous administration, distributing widely throughout the organism to target large or diffused biological structures. Specifically, researchers were interested in creating systemic capsids that could target the peripheral (PNS) and central (CNS) nervous systems. Targeting the CNS requires crossing the blood-brain barrier (BBB), and a systemic AAV capsid could avoid invasive methods such as intracranial injection. Systemic capsids are often derived from the AAV9 serotype, which is one of the few serotypes that can naturally cross the BBB. The Viviana Gradinaru lab has developed a number of systemic rAAV vectors that can target whole organs or specific cell types of the CNS (e.g. AAV-PHP.eB) and the PNS (e.g. AAV-PHP.S) in rodents and non-human primates. Read our blog post on systemic capsids for more information.
Browse Addgene’s in-stock viral preps.
AAV retrograde (AAVrg)
Directed evolution has also been employed to engineer an rAAV serotype variant able to display neuronal retrograde transport, AAVrg. Retrograde transport refers to the direction of movement within a neuron from the terminal part of the axon towards the soma, or main body of the cell. Therefore, when injected in a specific brain region, AAVrg would enter and track the terminal axons present in that region and move towards the soma of the cell, which may be in another region. In other words, AAVrg allows you to track neurons that send information to a given region. This allows researchers to trace connections between neurons, map the brain, and better understand brain structure.
Read our blog post on AAVrg for more information.
Self-complementary AAV (scAAV)
One downside of the use of AAVs as viral vectors is their single-stranded DNA genome. Because the rAAV vector depends on the cell’s DNA replication machinery to synthesize the complementary strand, transgene expression may be delayed. To overcome this rate-limiting step, scAAVs contain a dimeric inverted repeat DNA molecule that spontaneously reanneals upon infection, forming a double-stranded DNA (dsDNA) template. This eliminates the requirement for host cell DNA synthesis but further limits the packaging capacity of rAAV vectors to ~2.4 kb.
Common Uses of AAV Vectors
rAAV vectors are valuable in diverse applications due to their unique properties, including their non-pathogenic nature, ability to target specific tissues and cells, and capacity for long-term gene expression. These features make them ideal for gene therapy, neuroscience research, and other biomedical fields.
Gene Therapy
One of the major applications of rAAV vectors is in the field of gene therapy to treat genetic disorders. rAAV vectors are favored over other viral delivery systems due to their non-pathogenicity, ability to efficiently deliver genes to a wide range of tissues with minimal immune response, and the low risk of insertional mutagenesis. Several AAV-based gene therapies have been approved by the US Food and Drug Administration (FDA) to treat a diversity of diseases such as inherited retinal disease, spinal muscular atrophy, hemophilia A and B, and Duchenne muscular dystrophy. Many clinical trials of AAV-based gene therapies are underway and are promising for the treatment of other major diseases such as cystic fibrosis and cancer.
Neuronal Tracing
Recombinant AAV vectors have been widely used by neuroscientists to study neural connections and brain structure due to their small size, low immunogenicity, and limited ability to spread after injection. Different AAV serotypes exhibit strong neuronal tropism, meaning they can efficiently target specific brain regions or types of neurons, and are capable of both anterograde (from soma to axon) and retrograde (from axon to soma) transport. Many rAAV vectors have been engineered to express fluorescent proteins, often under strong neuron-specific promoters or neural Cre-dependent expression, to visualize connections between neurons within the brain. Large-scale examples of mapping the brain through neuronal tracing with rAAV vectors include the Allen Mouse Brain Connectivity Atlas and the Janelia Campus MouseLight project . rAAV vectors have also been developed for their use with rabies virus (RABV) in monosynaptic retrograde tracing of neurons. In this system, the rAAV vector is used prior to RABV infection to pre-label specific neurons for infection and to express a viral element necessary for RABV trans-synaptic transport. Learn more about these and other initiatives in our blog post on using AAV for neuronal tracing.
Browse AAV vectors for monosynaptic neuronal tracing available at Addgene.
Optogenetics
In optogenetics, microbial opsins and other optical tools developed by researchers are used to detect, measure, and control molecular signals of cells (commonly neurons) in response to light. The short coding sequences of channelrhodopsins, halorhodopsins, and other optogenetic genes makes them suitable to be packaged in the relatively small rAAV capsids. Learn more about optogenetics by watching our Optogenetics 101 video or by visiting Addgene's Optogenetics Guide.
Browse optogenetics plasmids available at Addgene.
CRISPR/Cas9 Delivery
rAAV vectors are a popular choice for the delivery of CRISPR/Cas9 elements, making it one of the most common methods for in vivo CRISPR-based genome editing. Packaging the Cas9 from Streptococcus pyogenes (SpCas9) and a gRNA together into an rAAV capsid is challenging due to its limited packaging capacity, especially if any additional regulatory elements are needed. One strategy to overcome this obstacle, has been to use smaller Cas9 orthologs, such as SaCas9 from Staphylococcus aureus. The Feng Zhang lab has developed plasmids containing SaCas9 optimized for their use in rAAV vectors. Read our blog post on overcoming AAV size limitation for CRISPR delivery for more information.
Split AAVs to increase packaging capacity
Another strategy to deliver cargo too large to fit within the rAAV capsid, such as Cas9-derived plasmids and base/prime editors, is to create split AAVs. Here, a longer transgene is broken down between two AAV transfer plasmids and packaged in separate rAAV vectors which are then co-delivered to the host cells. The full-length transgene is later reconstructed within the cell via different mechanisms. One of these methods uses splice site donor and acceptor sequences to reconstitute the two pieces of the transgene. The first vector contains the 5' portion of the gene and a splice donor sequence at its 3' end, while the second vector carries the 3' portion of the gene with a splice acceptor sequence at its 5' end. When these viral vectors co-infect a cell, they form concatemers and splice together, reconstructing the full-length transgene that can then be expressed. This method allows for delivery of larger transgenes, but expression is still much less efficient than with a single rAAV vector (∼5%). Gene fragments can also be reassembled via homologous recombination, when both sequences present substantial sequence overlap. Co-expression induces homologous recombination and expression of the full-length transgene, but also at a very low efficiency (less than 1% of wild-type). Another split AAV method which has shown higher efficiency in delivering large genes makes use of split inteins, proteins that carry out a naturally occurring process known as protein trans-splicing. As these methods are made more efficient, the use of rAAV vectors will no longer be limited to small transgenes, enabling the development of further AAV applications.
Read our blog post on split AAVs for more information.
Viral Vector Safety
AAVs are replication incompetent and not known to cause disease in humans. Most, if not all of Addgene’s transfer plasmids are replication deficient. These plasmids can be used to create viral vectors capable of infecting target cells, but cannot produce any new viral particles after the initial infection. The risk of spontaneously occurring replication-competent rAAV vectors is extremely reduced by dividing the viral genome between the transfer and the helper plasmid, and by providing the helper E1 gene in trans within the packaging cell line.
rAAV vectors present low oncogenic potential because they do not integrate in the human chromosome, which reduces the risk of insertional mutagenesis. However, random integration events can occur at a very low frequency.
As with most experiments, infection risks occur with contact to mucous membranes or broken skin. Needle sticks and ripped gloves are common points of entry. Biosafety should always be considered with respect to the precise nature of experiments being performed. Your biosafety office can provide more information on your institution's best practices with regard to AAV-related research.
See Addgene’s Biosafety Resource Guide for more information and resources on viral safety.
Resources and References
Addgene's Viral Vector Packaging Services
Resources on addgene.org
Resources on Addgene's Blog
- Parts of the AAV Transfer Plasmid
- Parts of the AAV Packaging Plasmid
- Producing Your rAAV
- AAV Serotypes and Tissue Tropism
- Adeno Associated Virus (AAV) for Cell and Gene Therapy
- Adeno-associated Viruses (AAVs) for Genome Editing
- Using AAV for Neuronal Tracing
- More Viral Vectors 101 topics
- More AAV Vector topics
Addgene Protocols
References
Burger, C., Gorbatyuk, O. S., Velardo, M. J., Peden, C. S., Williams, P., Zolotukhin, S., Reier, P. J., Mandel, R. J., & Muzyczka, N. (2004). Recombinant AAV viral vectors pseudotyped with viral capsids from serotypes 1, 2, and 5 display differential efficiency and cell tropism after delivery to different regions of the central nervous system. Molecular Therapy, 10(2), 302–317. https://doi.org/10.1016/j.ymthe.2004.05.024 (Link opens in a new window) PMID: 15294177 (Link opens in a new window)
Chan, K. Y., Jang, M. J., Yoo, B. B., Greenbaum, A., Ravi, N., Wu, W. L., Sánchez-Guardado, L., Lois, C., Mazmanian, S. K., Deverman, B. E., & Gradinaru, V. (2017). Engineered AAVs for efficient noninvasive gene delivery to the central and peripheral nervous systems. Nature Neuroscience, 20(8), 1172–1179. https://doi.org/10.1038/nn.4593 (Link opens in a new window) PMID: 28671695 (Link opens in a new window)
Deyle, D. R., & Russell, D. W. (2009). Adeno-associated virus vector integration. Current Opinion in Molecular Therapeutics, 11(4), 442–447. PMID: 19649989 (Link opens in a new window)
Duan, D., Yue, Y., & Engelhardt, J. F. (2001). Expanding AAV packaging capacity with trans-splicing or overlapping vectors: a quantitative comparison. Molecular Therapy, 4(4), 383–391. https://doi.org/10.1006/mthe.2001.0456 (Link opens in a new window) PMID: 11592843 (Link opens in a new window)
Grimm, D., & Kay, M. A. (2003). From virus evolution to vector revolution: use of naturally occurring serotypes of adeno-associated virus (AAV) as novel vectors for human gene therapy. Current Gene Therapy, 3(4), 281–304. https://doi.org/10.2174/1566523034578285 (Link opens in a new window) PMID: 12871018 (Link opens in a new window)
Hüser, D., Weger, S., & Heilbronn, R. (2003). Packaging of human chromosome 19-specific adeno-associated virus (AAV) integration sites in AAV virions during AAV wild-type and recombinant AAV vector production. Journal of Virology, 77(8), 4881–4887. https://doi.org/10.1128/jvi.77.8.4881-4887.2003 (Link opens in a new window) PMID: 12663794 (Link opens in a new window)
Kwon, I., & Schaffer, D. V. (2008). Designer gene delivery vectors: molecular engineering and evolution of adeno-associated viral vectors for enhanced gene transfer. Pharmaceutical Research, 25(3), 489–499. https://doi.org/10.1007/s11095-007-9431-0 (Link opens in a new window) PMID: 17763830 (Link opens in a new window)
Liu, D., Li, T., Liu, L., Che, X., Li, X., Liu, C., & Wu, G. (2024). Adeno-associated virus therapies: pioneering solutions for human genetic diseases. Cytokine & Growth Factor Reviews, 80, 109–120. https://doi.org/10.1016/j.cytogfr.2024.09.003 (Link opens in a new window) PMID: 39322487 (Link opens in a new window)
Matsushita, T., Okada, T., Inaba, T., Mizukami, H., Ozawa, K., & Colosi, P. (2004). The adenovirus E1A and E1B19K genes provide a helper function for transfection-based adeno-associated virus vector production. Journal of General Virology, 85(Pt 8), 2209–2214. https://doi.org/10.1099/vir.0.79940-0 (Link opens in a new window) PMID: 15269360 (Link opens in a new window)
McCarty, D. M., Monahan, P. E., & Samulski, R. J. (2001). Self-complementary recombinant adeno-associated virus (scAAV) vectors promote efficient transduction independently of DNA synthesis. Gene Therapy, 8(16), 1248–1254. https://doi.org/10.1038/sj.gt.3301514 (Link opens in a new window) PMID: 11509958 (Link opens in a new window)
Merten, O. W. (2024). Development of stable packaging and producer cell lines for the production of AAV vectors. Microorganisms, 12(2), 384. https://doi.org/10.3390/microorganisms12020384 (Link opens in a new window) PMID: 38399788 (Link opens in a new window)
Naso, M. F., Tomkowicz, B., Perry, W. L. 3rd, & Strohl, W. R. (2017). Adeno-associated virus (AAV) as a vector for gene therapy. BioDrugs, 31(4), 317–334. https://doi.org/10.1007/s40259-017-0234-5 (Link opens in a new window) PMID: 28669112 (Link opens in a new window)
Tervo, D. G., Hwang, B. Y., Viswanathan, S., Gaj, T., Lavzin, M., Ritola, K. D., Lindo, S., Michael, S., Kuleshova, E., Ojala, D., Huang, C. C., Gerfen, C. R., Schiller, J., Dudman, J. T., Hantman, A. W., Looger, L. L., Schaffer, D. V., & Karpova, A. Y. (2016). A designer AAV variant permits efficient retrograde access to projection neurons. Neuron, 92(2), 372–382. https://doi.org/10.1016/j.neuron.2016.09.021 (Link opens in a new window) PMID: 27720486 (Link opens in a new window)
Wu, Z., Asokan, A., & Samulski, R. J. (2006). Adeno-associated virus serotypes: vector toolkit for human gene therapy. Molecular Therapy, 14(3), 316–327. https://doi.org/10.1016/j.ymthe.2006.05.009 (Link opens in a new window) PMID: 16824801 (Link opens in a new window)
AAV Plasmid Elements
| Plasmid Type | Element | Delivery relative to transgene | Purpose |
|---|---|---|---|
| Transfer/Shuttle plasmid | 5' ITR (LITR) | in cis | Left Inverted Terminal Repeat, contains a Rep binding element (RBE) that serves as a binding site for Rep proteins essential for viral DNA replication and packaging. |
| 3' ITR (RITR) | in cis | Right Inverted Terminal Repeat, contains a Rep binding element (RBE) that serves as a binding site for Rep proteins essential for viral DNA replication and packaging. | |
| WPRE | in cis | Woodchuck hepatitis virus post‐transcriptional regulatory element; stimulates the expression of transgenes via increased nuclear export. | |
| Poly(A) signal | in cis | The polyadenylation or poly(A) signal aids in the nuclear export of RNA and RNA translation, and promotes RNA transcript longevity. Located between WPRE and 3'ITR. Not required but often included. | |
| Packaging plasmid | Rep | in trans | The rep gene encodes four proteins that are required for viral genome replication and packaging: Rep78, Rep68, Rep52, and Rep40. |
| Cap | in trans | The cap gene encodes three AAV viral capsid proteins VP1, VP2, and VP3, which together form the outer capsid shell that protects the viral genome. | |
| Helper plasmid | E4 | in trans | Adenoviral early gene that encodes for proteins essential for AAV genome replication and packaging. |
| E2A | in trans | Adenoviral early gene that encodes a DNA-binding protein (DBP) that enhances DNA replication by stimulating elongation and stabilizing the displaced single-stranded DNA. | |
| VA | in trans | Adenoviral gene that encodes for virus-associated RNAs involved in regulating translation and inhibiting cellular anti-viral defenses. |
Glossary
| Term | Definition |
|---|---|
| Adeno-associated virus (AAV) | Wildtype, small, nonenveloped viruses from the Parvoviridae family. |
| Anterograde transport | Movement of molecules, vesicles or organelles from the main cell body, or soma, towards the synapse at the axons. |
| Blood-brain barrier (BBB) | Layer of cells that prevent the entry of toxins and pathogens from the blood to the CNS. |
| Concatemer | A DNA segment composed of multiple, repeated DNA sequences linked end-to-end. |
| Episome | Genetic element inside the cell that remains as a part of the eukaryotic genome without integration and can replicate independently. |
| Genetic cargo | DNA introduced to cells within the viral vector capsid. |
| Icosahedral | Having the shape of a polyhedron with 20 faces, each being an equilateral triangle. |
| Immunogenicity | The ability of a molecule or substance to induce an immune response in the body. |
| in cis | In the context of viral vector production, in cis refers to genetic elements located in the same plasmid as the gene of interest. |
| in trans | In the context of viral vector production, in trans refers to genetic elements provided outside of the plasmid containing the gene of interest (i.e. other plasmids, packaging cell line). |
| Infection | The natural process of entry and multiplication of a pathogen, like viruses or bacteria, within a host organism, leading to disease. |
| Insertional mutagenesis | Random insertion of DNA elements into the genome, often altering gene activity. |
| Latent infection | A state where a pathogen is present in an organism but not actively causing symptoms or replicating. |
| Lytic cycle | Viral replication process where the virus uses the cell machinery to produce new viruses, eventually lysing the cell to release them. |
| Optogenetics | Biological technique to control the activity of cells, often neurons, with light. |
| Pseudotyping | The production of viral vectors (or viruses) using viral envelope proteins from different species. Used to alter infectivity and tropism. |
| Recombinant adeno-associated viral (rAAV) vector | Modified form of AAV used to deliver genetic material into cells. |
| Replication-competent rAAV | Mutated rAAV vector that has the ability to replicate and reproduce within host cells as a result of a spontaneous crossover event between the rAAV vector and the adenoviral genes provided in trans. |
| Retrograde transport | Movement of molecules, vesicles or organelles from the synapse at the axons towards the main cell body, or soma. |
| Transduction | The process of artificially introducing foreign DNA into eukaryotic cells using viral vectors. |
| Transfection | The process of artificially introducing foreign DNA into eukaryotic cells using non-viral methods. |
| Transformation | The process where bacteria take up DNA from their environment. |
| Tropism | The specific types of cells and tissues that a virus can infect and replicate in. |
Credits
- Contributing Authors
- Written and reviewed by the Scientific Curation team at Addgene.
- Media Credits
- Figures created with BioRender (Link opens in a new window)
- Last Updated
- Content last reviewed on 9 June 2025.